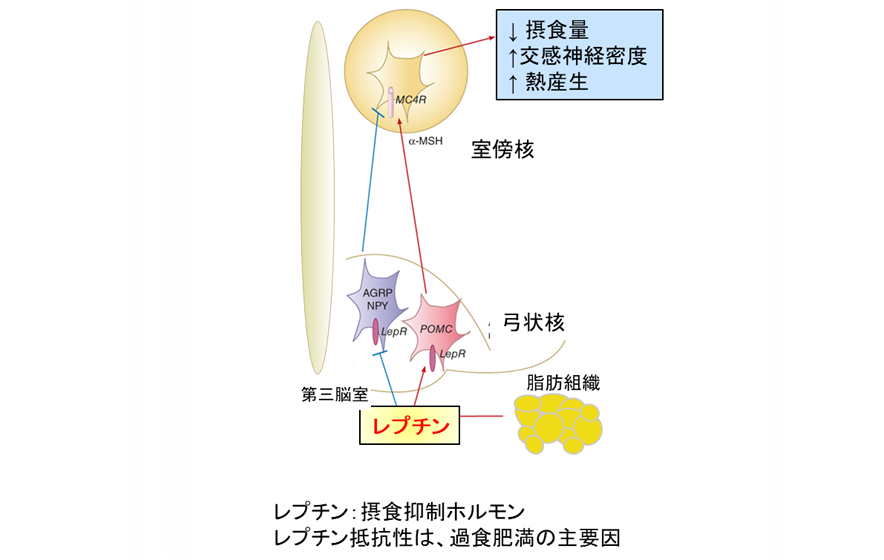
栄養刺激に対しては、遺伝子改変マウスにのみ、弓状核(視床下部の内側に存在する恒常性摂食調節機構を制御する領域)にある活性化したニューロン数の減少が確認できました。弓状核は食欲抑制の働きを持つ場所です。
また、中枢神経細胞特異的にヒストン脱メチル化酵素を持たないマウスを作成したところ、摂食増加、脂肪組織の交感神経密度の低下を伴い、エネルギー消費が低下することもわかってきました。
 ヒストン脱メチル化の異常はレプチン抵抗性を示し、摂食行動の抑制が効かなくなる
ヒストン脱メチル化の異常はレプチン抵抗性を示し、摂食行動の抑制が効かなくなる
東北大学
酒井 寿郎教授
Juro Sakai
栄養刺激に対しては、遺伝子改変マウスにのみ、弓状核(視床下部の内側に存在する恒常性摂食調節機構を制御する領域)にある活性化したニューロン数の減少が確認できました。弓状核は食欲抑制の働きを持つ場所です。
また、中枢神経細胞特異的にヒストン脱メチル化酵素を持たないマウスを作成したところ、摂食増加、脂肪組織の交感神経密度の低下を伴い、エネルギー消費が低下することもわかってきました。
ヒストン脱メチル化の異常はレプチン抵抗性を示し、摂食行動の抑制が効かなくなる本研究では、寒冷刺激を感知したことでリン酸化したヒストン脱メチル化酵素のネガティブ調整因子である、脱リン酸化酵素複合体を発見しました。そして、この脱リン酸化酵素複合体がベージュ脂肪細胞の誘導に関与していることもわかりました。
未分化な細胞が白色脂肪細胞、褐色脂肪細胞、ベージュ脂肪細胞のいずれかへと分化するためには、3つのステップがあります。ステップ1:エピゲノムによる必要な遺伝子の選択、ステップ2:転写因子・転写共役因子による転写、ステップ3:RNA転写後修飾です。
このステップ1から2に対して脱リン酸化酵素複合体が環境刺激をインプットすることにより、エピゲノム経路と転写経路を統合させ、ベージュ脂肪細胞への誘導を制御していることが明らかになりました。
また、ステップ3がステップ1を非線形的に制御する新たな機構が存在すること、ステップ3が代謝性疾患と関連することも示唆されました。
これまで概念的理解に留まっていた「環境因子が疾患発症を制御する機構」が、DNA/ヒストン/RNAの化学修飾レベルで明らかになりつつあります。これを解明することで、環境刺激を模倣した分子介入による疾患の治療・予防が可能になると期待できます。
私の専門分野は内分泌系だったため、神経系はあまり明るくありませんでした。しかし、レプチン抵抗性には神経回路を介した制御が存在すると考えていたため、末梢から中枢までを本格的に勉強する必要がありました。
大変でしたが、この一般研究助成に採択していただいたおかげで、必要な知識と技術をいくつも習得し、共同研究が実現して、セットアップを進めることができたのです。
東京大学の谷内江望先生や、九州大学の小川佳宏先生、名古屋大学の菅波孝祥先生など、同じ研究分野で知り合いの先生方が、セコム科学技術振興財団の研究助成に採択されていました。チャレンジングな研究を発展させてくれる制度だと感じ、以前から気になっていました。
この研究は短期間で成果を出すことはできませんが、多くのファンドは助成期間が1〜2年しかありません。長期間サポートしてくださるのはセコム財団だけでしたので、思い切って申請に踏み切りました。
ただ、10年近く温めていたものの、当時はまだ仮説の域を出ないアイデアでした。申請した時は自信がなかったのですが、採択していただき、たいへん嬉しかったです。探索的な研究や仮説の検証に長い時間が必要でしたが、大きな助成金で長期間サポートしていただいたおかげで、できることがどんどん増えて、課題を深掘りすることができました。いくつもの達成感とともに、私のエピゲノム研究が大きく進展したのです。本当に感謝の言葉もありません。
 面談では厳しいご指摘もあり、鋭いところを突かれたりもしたため、採択通知が届いたときは非常に嬉しかった
面談では厳しいご指摘もあり、鋭いところを突かれたりもしたため、採択通知が届いたときは非常に嬉しかった