第1回でお話しいただいた、活性化YAP誘導性の異常細胞排除に必須の因子であるプロスタグランジンE2(PGE2)については、何かわかりましたか。
哺乳動物培養細胞を用いた、YAP依存性の異常細胞排除において、PGE2が必須であることを報告しました。PGE2がその受容体を介して細胞内シグナル伝達系を活性化し、細胞骨格の動態を制御することは突き止めました。隣接する正常細胞が異常細胞を排除するために必要な過程だと考えています。
しかしながら、PGE2を添加させても細胞排除は誘導されません。つまり、異常細胞を骨格変化させ、排除するためにはPGE2だけでは不十分であること、他の因子も必要であることが示唆されました。マウスの肝臓(in vivo)の異常細胞排除においても、PGE2添加だけでは異常細胞の類洞側への移動、細胞死誘導は誘導できず、同様に、他の因子の必要性が示唆されました。
PGE2は、異常細胞排除機構の必要条件であっても、十分条件にはならなかった、ということですか。
はい、そうです。傷害のシグナルが付加されたときに細胞骨格を変化させる新たな遺伝子が出現し、YAPはその遺伝子発現を誘導していることは確認できました。しかし、PGE2はこの遺伝子発現に関わっていませんでした。
異常細胞排除という現象は最近注目された細胞応答のため、まだ研究が進んでいません。「異常細胞の発生→異常細胞の感知→細胞間接着の解除→細胞排除」といった複数の過程を持つ興味深い現象です。これらの過程を解明するために、引き続き網羅的な遺伝子発現解析を含む実験を行っていきます。
PGE2以外の排除因子の調査や、ヒトの細胞を用いた実験系の構築については、いかがでしたか。
異常細胞排除を促進する化合物の同定は、最終目的である新しい治療法の実現に不可欠です。副作用の少ない新規化合物の同定を目指して、長崎大学が保有する海洋微生物抽出物ライブラリーと、北里大学が保有する天然化合物ライブラリーを使用し、スクリーニングを行っています。その結果、異常細胞排除を促進する複数の海産物のフラクション(成分)を同定しています。現在はそれをさらに分画し、それぞれを精製し、分子構造の同定を進めているところです。
また、肝臓オルガノイド製作用の材料は、コロナ禍の影響で入手困難な状態が続いているため、ヒト細胞を用いた異常細胞排除実験系の確立は遅れています。
一方、独自に進めていた老化マウスの製作には成功しました。このマウスを用いた実験によって、加齢が異常細胞排除に影響するのかどうかを検討しています。
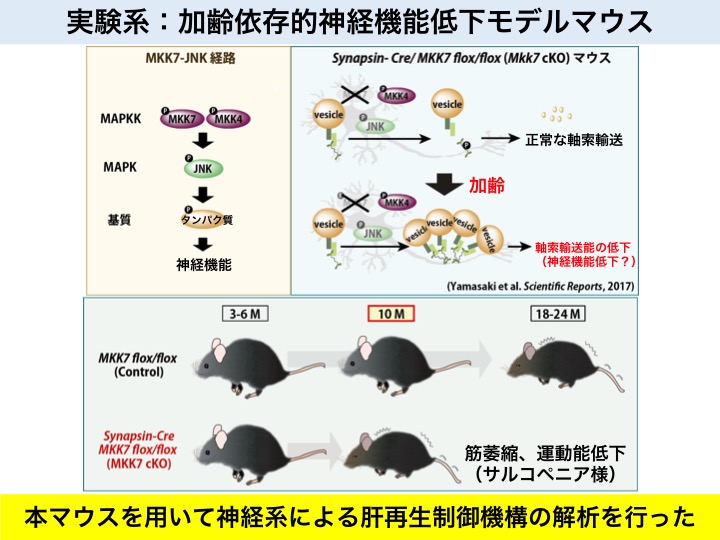
加齢によって異常細胞排除制御因子の機能低下が生じ、その結果、異常細胞排除の低下が生じる可能性が考えられる。そこで、その評価を手助けするために、独自の老化マウスの作出を行った
老化マウスとは、どのような特徴を持っているのでしょうか。
私たちの体は歳をとると、組織や臓器の機能が衰えます。興味深いことに、組織や臓器が一つずつバラバラに悪くなっていくのではなく、ほぼ同時進行的に低下していきます。この現象について、私たちは各組織や臓器の寿命が同時期に寿命を迎えるというより、システムを制御する神経機能の低下が全体に影響を及ぼしているのだと考えました。
神経細胞は、核を含む細胞体、軸索、樹状突起など複数の部位に分かれています。細胞体から離れた部位へのmRNAやタンパク質などの分子の供給は、軸索輸送を通して行われます。
一般的に、マウスは生後2年から脱毛や運動機能の低下といった老化現象が観察されますが、我々の10カ月齢の老化マウスは、軸索輸送能が低下し、筋萎縮の亢進や運動能の低下などの老化現象が観察されます。
老化マウスの肝臓を70%切除すると、6カ月齢では生存しましたが、10カ月齢では肝再生能の低下が原因で、死亡しました。
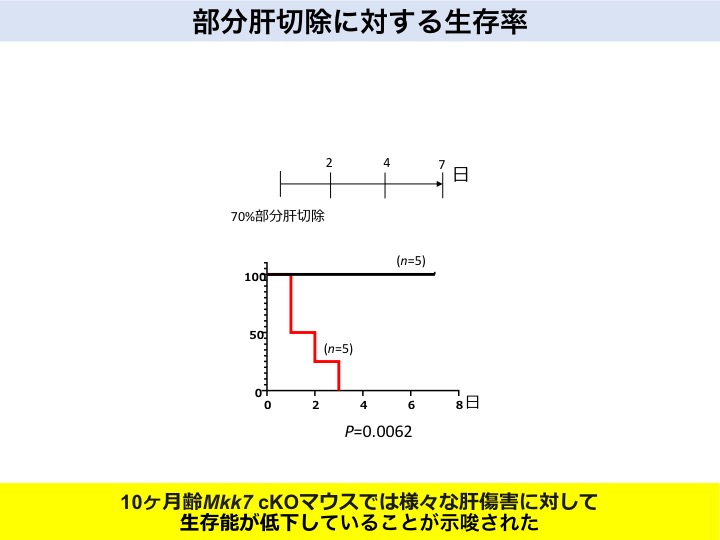
加齢依存的神経機能低下モデルマウス(老化マウス)を用いた、肝臓70%切除による生存率の実験
老化が始まったマウスは、肝臓の再生能力も低下するのですね。
はい、そうです。我々の老化マウスは、先天的に成熟神経細胞特異的にある遺伝子が欠損していますが、6カ月齢までの若いうちはとくに肝再生能に問題はありません。しかし10カ月齢になると通常よりも早く老化現象が生じ、肝再生能力の低下が起こります。つまり、老化によって神経機能が低下すると、肝臓の恒常性を維持する再生能力は低下することが示唆されました。
YAP依存の異常細胞排除機構にも、神経機能や老化が影響を与えるのか否か。もし本マウスを用いて、加齢依存的な細胞排除の低下が観察されれば、肝臓の恒常性の維持には神経機能の制御も重要な要素になるため、引き続き研究を進めていくつもりです。