はい。なお、MDRA感染がインフラマソーム活性化により悪化することから、その詳細な分子メカニズムを調べるために、caspase-11の下流シグナルを調べました。caspase-11は直接ガスダーミンDを活性化できるので、まずガスダーミンD欠損マウスにMDRAを感染させたところ、caspase-11欠損マウスと同様に臓器内菌数の低下が観察されました。また、caspase-11は二次的にNLRP3インフラマソームを活性化できるのでそちらの関与も検討したところ、NLRP3欠損マウスでは顕著な臓器内菌数の減少は認められませんでした。
以上の結果から、MDRA感染においては、細胞膜に孔を形成してパイロトーシスを引き起こすガスダーミンDによって細胞膜傷害が誘導されることにより、病態が悪化することが明らかになりました。
MRSA感染において、インフラマソーム応答の中心となるcaspase-11は、グラム陰性菌のLPSに対する細胞内受容体として知られています。そこで、LPSがインフラマソーム応答の誘導に関与している可能性を考えて、アシネトバクターのLPS合成酵素を欠損した変異株(LPS欠損株)をマクロファージに感染させ、野生株を感染させた場合と比較しました。その結果、野生株感染の場合に比べて、LPS欠損株ではIL-1βの産生量が激減したのに対し、インフラマソームに基づかないTNFαの産生量には、大きな変化が見られませんでした。
さらに、LPS合成酵素をプラスミド性に相補したところ、IL-1βの産生量が回復しました。このことから、LPSがアシネトバクターの主要なインフラマソームリガンドとなっていることが明らかになりました。
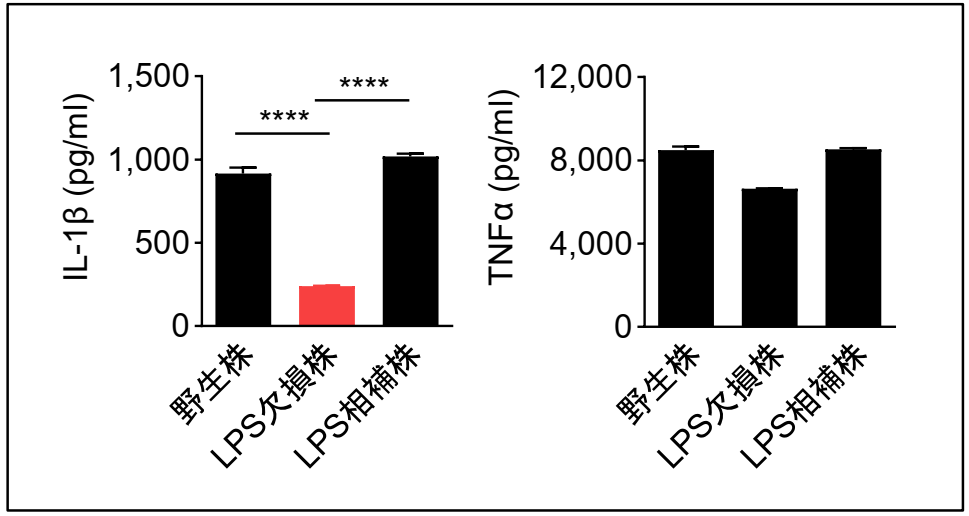
アシネトバクターの野生型とLPS欠損株、相補株をマクロファージに感染させた場合のIL-1βとTNFαの産生量
ここまでの結果をまとめると、黄色ブドウ球菌やリステリアなどのグラム陽性菌は、インフラマソームを活性化することで、caspase-1によって誘導された炎症性サイトカインのIL-18およびIL-1βが病態を悪化させます。
これに対し、アシネトバクターなどのグラム陰性菌は、LPSを介してインフラマソームを活性化します。そして、caspase-11がガスダーミンDを活性化することで細胞膜傷害が生じ、病態を悪化させるのです。グラム陰性菌感染の場合にも、細胞内受容体NLRP3が病原体関連分子パターン(PAMPs)や損傷関連分子パターン(DAMPs)を認識し、インフラマソームが活性化して、IL-18とIL-1βが産生されます。しかし、これらはほとんど感染病態には影響しないことがわかりました。
なお、グラム陽性菌感染において生じるcaspase-1依存のインフラマソームを「古典的インフラマソーム(canonical inflammasome)」、グラム陰性菌において生じるcaspase-11を中心としたインフラマソームを「非古典的インフラマソーム(non-canonical inflammasome)」と呼びます。
本格研究2年目までに得られた知見を踏まえたうえで、グラム陽性菌感染において、病理的なインフラマソーム応答の中核となるASC speck形成の詳しいメカニズムや、インフラマソーム応答以降のプロセスにおいて、具体的にどのような分子メカニズムで感染病態が悪化するのかを調べています。
また、もう一つのテーマである、炎症記憶に関する研究にも取り組んでいます。クロマチンの修飾やリモデリング、DNAのメチレーションによって炎症が細胞に記憶されることは以前から指摘されていますが、近年、細胞内受容体のAIM2やNLRP3が炎症記憶に関わっていることが報告されており、未知のメカニズムによりインフラマソーム炎症が個体に記憶されていると考えられます。
このように、インフラマソームは一過性の炎症応答だけでなく、その後の生体応答に、長期的に影響を及ぼしている可能性が高いのです。感染症は生涯に何度も患うケースが多いので、1回目の感染炎症がその後の感染に及ぼす影響を検討しているところです。