
「統合的プロテオミクスによる細胞内翻訳後修飾制御機構の多層的理解」
膜トポロジーの反転は膜タンパク質の新たな機能獲得様式
本研究では、GGCXの膜トポロジー反転というテーマにも取り組みました。
細胞膜には「膜タンパク質」が存在し、周囲の細胞との相互作用や細胞内外の物質や情報をやりとりするために、細胞膜上で一定の方向、すなわち細胞の外側か内側を向いています。この膜タンパク質の配向を「トポロジー」と呼んでいます。
GGCXは小胞体膜に存在する複数回膜貫通タンパク質で、基質と結合する酵素活性部位は、小胞体内腔側、つまり細胞にとっては細胞外側と同側にあるとこれまで考えられていました。つまり、GGCXのトポロジーは外側であるといえます。しかし、IPS-1は細胞内側に存在しています。通常であれば、GGCXはIPS-1などの細胞質側のタンパク質を修飾することはできません。
では、どのように修飾しているのか。そのメカニズムについて調べた結果、ウイルス感染に応答する形でGGCXが膜トポロジーを反転させ、酵素活性部位が細胞質側を向くことで、IPS-1がカルボキシル化修飾されることがわかりました。
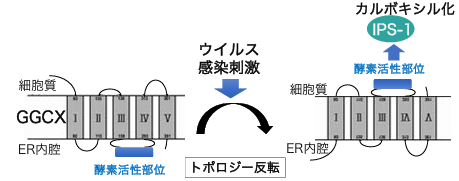 ウイルス感染に応答してGGCXの膜トポロジーが反転。酵素活性部位が細胞質側を向き、細胞内タンパク質のカルボキシル化修飾が可能となる
ウイルス感染に応答してGGCXの膜トポロジーが反転。酵素活性部位が細胞質側を向き、細胞内タンパク質のカルボキシル化修飾が可能となる複数回膜貫通タンパク質が刺激に応答してトポロジー反転を起こすという現象自体は、すでに10年前に発見されましたが、現在でもGGCXを含め3つしか報告されておらず、例がまだほとんどありません。更に最新の研究では、膜タンパク質のトポロジー反転は単なるエラー現象であり、校正システム(タンパク質の過配送をやり直す仕組み)によって膜の挿入のやり直しや分解を受けることが報告されています。
しかしながら、GGCXの膜トポロジー反転は、ウイルス感染に応答して、細胞内タンパク質を翻訳後修飾するという合理的な目的で起こっています。このことから、GGCXの膜トポロジー反転は単なるエラーではなく、ウイルス感染などのストレス下において膜タンパク質が新しい機能を獲得するための様式であり、校正システムからエスケープされているのではないかと考えました。
なお、本研究では、こうした成果に基づいて膜トポロジー反転の制御機構の解明に取り組むとともに、GGCX以外にトポロジー反転制御を受ける膜タンパク質の網羅的探索と候補因子の機能解析を行いました。将来的には、膜トポロジー反転を人為的に操作することで、どのような影響が生じるのかも、調べたいと思っています。
基礎生物学の立場から、生命現象や疾患発症のメカニズム解明を目指す
タンパク質がDNAからRNAを経て翻訳された後に、細胞内でどのように制御されるかはいまだ不明な点が多く、特に細胞内カルボキシル化修飾や膜トポロジー反転に関しては、研究の余地が残されています。生命現象や疾患発症のメカニズムを理解するうえで非常に重要な要素のため、ライフワークとして検証に取り組み、これを足掛かりに研究を発展させていく所存です。
また、複数の翻訳後修飾の組み合わせが細胞にどのような機能的変化をもたらすのかなど、翻訳語修飾を網羅的に調べる統合的プロテオミクスの手法の確立にも取り組みたいと考えています。そして、基礎生物学の立場から生物や生命現象の仕組みを解明し、新しいウイルス防御戦略や疾患治療法が提案できるよう研究を進めてまいります。
 本研究では、細胞内カルボキシル化制御に関わる候補因子も発見。これらの因子が、抗ウイルス応答を変化させるなどの機能を持つことが明らかになりつつある
本研究では、細胞内カルボキシル化制御に関わる候補因子も発見。これらの因子が、抗ウイルス応答を変化させるなどの機能を持つことが明らかになりつつある独立したばかりの若手研究者を手厚くサポートしてくれる助成
挑戦的研究助成については、以前所属していた研究室の先輩から教えてもらいました。年齢制限ぎりぎりでの応募でしたが、それよりも早い段階では業績が不足していたため、ちょうどよいタイミングだったと思います。
当時は独立したばかりで、しかも新設される研究室への赴任だったため、すべてを一から準備しなくてはならず、資金的に心もとない状態でした。そのような時に手厚くサポートしていただき、深く感謝しています。
特にありがたいのは、建設的な助言をいただけるメンタリングです。研究を発展させるためにはどうすればよいかなど、率直に温かいアドバイスが受けられるので、たいへん助かっています。
若手研究者の中には、独立したばかりで、経済的に苦しかったり、研究者としての方向性に悩んでいる方も多いかと思います。そのような時に強力なサポートとなり、またリスクを恐れない姿勢が高く評価される助成なので、ぜひチャレンジしてほしいと思います。
 独立したばかりの頃は、研究の方向性が正しいのか不安になりがちだった。メンタリングで一流の先生から丁寧な助言をいただき、たいへん参考になった
独立したばかりの頃は、研究の方向性が正しいのか不安になりがちだった。メンタリングで一流の先生から丁寧な助言をいただき、たいへん参考になった