

- 2009年3月
- 東京大学 大学院 理学系研究科 博士課程修了 博士(理学)
- 2009年4月
- ペンシルバニア大学 生物学部 研究員
- 2014年4月
- ペンシルバニア大学 生物学部 日本学術振興会 海外特別研究員
- 2015年4月
- 奈良先端科学技術大学院大学 助教(元さきがけ兼任研究員)
- 2022年4月
- 奈良先端科学技術大学 准教授
大学3年生で初めて植物の遺伝子研究に関心を持った
植物生理学に関心を持つようになったのは、大学3年生のときです。ちょうどその頃、シロイヌナズナのゲノム解読が完了したことを知り、植物の遺伝子研究は発展が見込めそうだと考えてこの分野を専攻しました。
そして大学院進学後、生物化学セミナーでエピジェネティクスの発表を聞いたことをきっかけに、エピゲノムが遺伝子の発現を促進・抑制する仕組みに強い興味を抱きました。現在は、エピジェネティクスの観点から植物の形態形成や、環境応答のメカニズムの解明に取り組んでいます。
 形態形成と応答変化は、ともすれば異なるテーマとして扱われがちだが、どちらもエピゲノムと深く関わる生理現象である
形態形成と応答変化は、ともすれば異なるテーマとして扱われがちだが、どちらもエピゲノムと深く関わる生理現象である植物の形態形成を制御する遺伝子や、遺伝子発現を制御するエピゲノムについて研究
植物の形態形成は、個体に因らず、ほぼ同一のパターンで進行します。まずは茎の先端部分にある分裂組織から葉が作られ、次に枝が伸びて、最後に茎が伸長しながら花が形成されるのです。このように同一のプロセスをたどるのは「形態形成を制御する遺伝子」の発現する順番が決まっているためと考えられます。
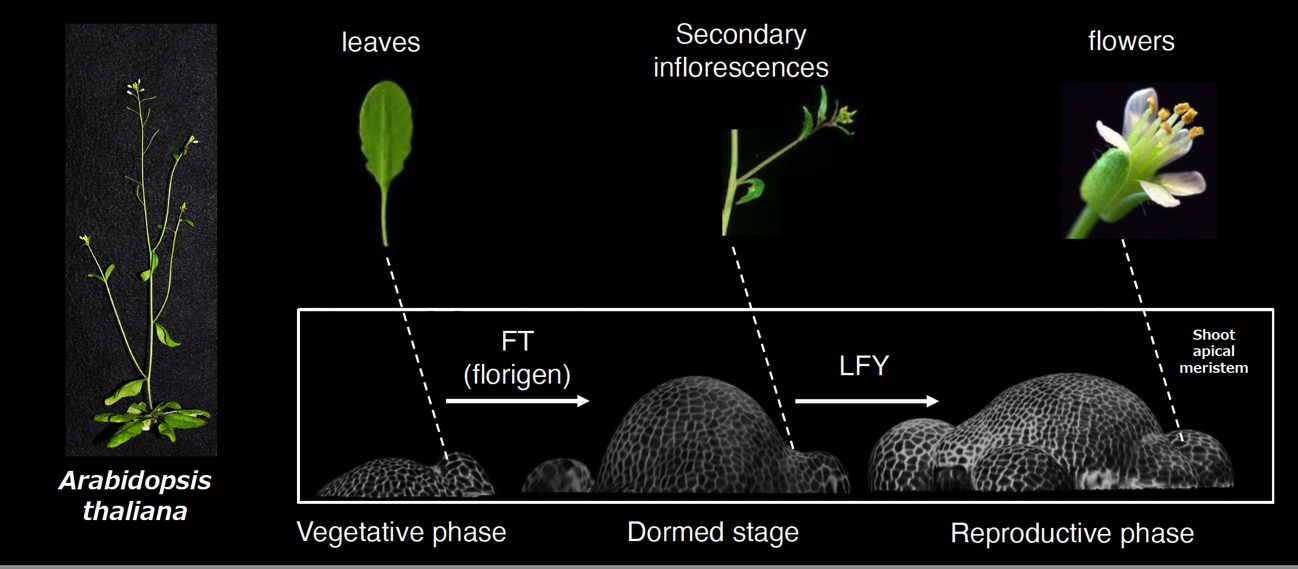 葉の形成から枝の形成に切り替える際にはFT遺伝子、枝の形成から花の形成に切り替える際にはLFY遺伝子というように、形態形成の各段階においてその現象を制御する遺伝子が存在する
葉の形成から枝の形成に切り替える際にはFT遺伝子、枝の形成から花の形成に切り替える際にはLFY遺伝子というように、形態形成の各段階においてその現象を制御する遺伝子が存在するそこで私は、植物の形態形成を制御する遺伝子や、その遺伝子が発現する順番、また形態形成に関わる遺伝子のネットワーク、そして遺伝子の発現を制御するエピゲノムに関心を持ち、研究に取り組んできました。たとえば、形態形成のメカニズムを確認するために、花を作るときに必要なLFY遺伝子の働きを強化することで、形態形成のパターンを変更して、枝形成の段階をスキップして即座に花の形成を引き起こすことに成功しています。
植物の高温順化が、ヒストン修飾の変化によって実現することを確認
動物や植物には、周囲の環境変化を感知して対応する「環境応答」という性質が備わっています。私はその中でも「高温順化」という現象に着目し、植物が高温ストレスにどのように適応するのか、またエピゲノムがどのような役割を果たしているのかについて研究してきました。
「高温順化」とは、植物に事前に中程度の高温を経験させると、強い高温ストレス下での生存が可能になるという現象です。実験と解析の結果、中程度の高温を経験した植物は、高温への耐性を獲得することがわかりました。
これは、ヒートショックプロテイン(熱ストレスによって損傷した細胞を、ストレスがかかる前の状態に修復する働きを持つタンパク質)の産生に関わる遺伝子発現を抑制するエピゲノム──ヒストン修飾のメチル基が、中程度の高温を経験することで除去され、短時間でヒートショックプロテインが増加したためと考えられます。ヒストン修飾とは遺伝子発現制御に関わる化学修飾であり、アセチル化、メチル化、リン酸化、ユビキチン化の4種類があります。
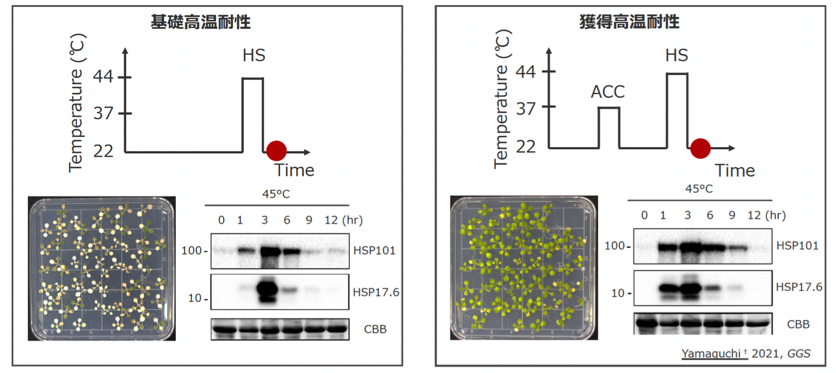 強い高温ストレスのみの場合(左)と、事前に中程度の高温を経験させた場合(右)
強い高温ストレスのみの場合(左)と、事前に中程度の高温を経験させた場合(右)DNAのメチル化や塩基配列の変更による細胞の性質変化は永続的で、次世代にも継承されます。しかし、高温ストレスが一時的なもので、迅速な対処が必要な場合は、植物はヒストン修飾やクロマチンの構造を変化させることで高温順化し、環境適応を実現しているのです。